Содержание страницы
- 1. Бактерии
- 1.1. Спорообразование
- 1.2. Размножение бактерий
- 1.3. Систематика бактерий
- 1.4. Дыхание бактерий
- 1.5. Молочнокислое брожение
- 1.6. Гетероферментативное молочнокислое брожение
- 1.7. Молочнокислые бактерии
- 1.8. Спиртовое брожение
- 1.9. Формы брожения по Нейбергу
- 1.10. Маслянокислое брожение
- 1.11. Маслянокислые бактерии
- 2. Грибы
- 3. Дрожжи
- 4. Вирусы
1. Бактерии
Бактерии представляют собой наиболее изученную группу микроорганизмов. Величина их – 0,4–10 мкм.
По форме бактерии подразделяют на несколько групп. Основными являются следующие:
- кокки – шаровидной формы, палочки (бактерии, бациллы);
- вибрионы – в виде запятых;
- спириллы – веретенообразной, слегка изогнутой формы и спирохеты – длинные, тонкие, сильно извитые (рис. 1).
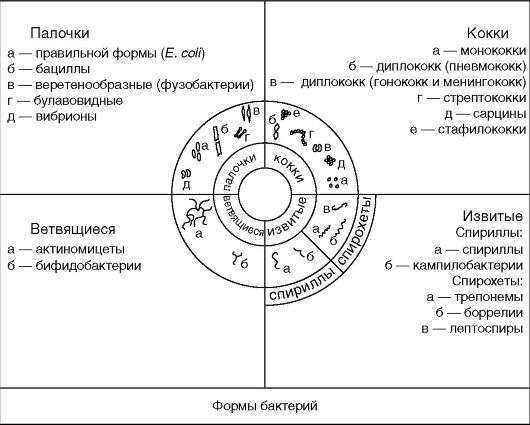
Рисунок 1 – Формы бактерий
Размеры и форма тела бактерий могут значительно изменяться под влиянием различных факторов внешней среды. Нетипичные, уродливые формы могут возникать под влиянием кислот, щелочей, температуры, накопления в среде продуктов жизнедеятельности и др.
Строение бактерий. Бактерии представлены лишь одной клеткой, которая является самостоятельным организмом (рис. 2).
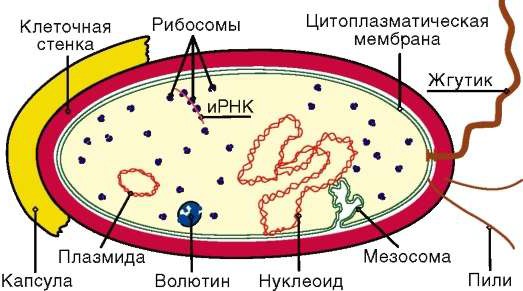
Рисунок 2 – Строение бактериальной клетки
Клетка бактерий покрыта оболочкой, которая выполняет защитные функции, придает клетке постоянную, характерную для нее форму (кокка, палочки, спириллы и др.). Она обладает свойством полупроницаемости: через нее питательные вещества проникают в клетку, а продукты жизнедеятельности клетки (продукты обмена) выходят в окружающую среду. Это относится к веществам, находящимся в сильно диспергированном виде в водных растворах (в состоянии истинных растворов). Крупные же молекулы, с большим молекулярным весом, через оболочку не проходят. Эта функция регулятора обмена веществ присуща всей оболочке, но в большей мере зависит от цитоплазматической мембраны.
Наружный, рыхлый слой оболочки у некоторых бактерий может ослизняться, образуя капсулу. Толщина капсул может во много раз превосходить диаметр клеток. Капсулы служат защитным покровом, участвуют в регуляции водного обмена, защищая клетки от высыхания. Состоят капсулы в основном из полисахаридов, гликопротеидов. Слизеобразующие бактерии, быстро размножаясь на поверхности субстратов, вызывают их порчу, а жидкие среды могут превращать в сплошную слизистую массу. Это явление иногда наблюдается в молоке, пиве, сахаристых экстрактах из свеклы и др. Слизеобразование активнее происходит при пониженных температурах – от 10 до минус 2 °С.
Цитоплазма – полужидкая, прозрачная масса белкового характера, которая является основной частью клетки. Наружная, более плотная часть цитоплазмы – цитоплазматическая мембрана – наряду с оболочкой участвует в регуляции обмена веществ с внешней средой. Во внутренней части цитоплазмы находятся важные клеточные структуры – рибосомы, мезосомы, ядро, запасные питательные вещества и др.
Рибосомы – зернистые образования, расположенные во всей цитоплазме. В них осуществляется синтез клеточных белков из поступающих веществ.
Мезосомы – тельца различной формы, находящиеся в цитоплазме и в пограничном с оболочкой слое. В них протекают энергетические процессы – освобождение энергии в результате окисления органических веществ пищи.
Ядро бактерий в отличие от других одноклеточных организмов некомпактно. Ядерное вещество равномерно распределено по всей цитоплазме. О наличии ядерного вещества судят по присутствию в составе бактериальной клетки дезоксирибонуклеиновых кислот (ДНК). ДНК является носителем наследственных свойств клетки. Ядро ответственно за передачу всех признаков родительских организмов потомству (форма, типичные размеры, физиологические свойства и др.). При размножении каждая вновь образуемая клетка бактерий получает полный набор нуклеиновых кислот, имеющихся у родительского организма. Свойства организма зашифрованы в структурных особенностях ДНК.
Запасные питательные вещества в виде гранул или капелек находятся в цитоплазме. Эти вещества накапливаются при благоприятных условиях и расходуются на дыхание, а также для построения различных структур тела клеток. Гранулы могут быть представлены крахмалом, гликогеном и белком волютином. Запасной жир образует мелкие шарообразные капли.
Жгутики представляют собой нитевидные образования, выступающие из-под цитоплазматической мембраны над поверхностью клетки. Жгутики являются органами движения. Расположение их может быть одиночным, в виде пучка на одном или обоих концах клетки и по всей поверхности. Жгутики очень тонки и легко теряются клетками при механических воздействиях, а также с возрастом.
Наличие жгутиков характерно не для всех бактерий, а лишь для некоторых палочковидных и шаровидных. Бактерии извитой формы передвигаются путем волнообразного изгибания тела.
1.1. Спорообразование
Некоторые бактерии обладают способностью образовывать споры. Это относится, прежде всего, к палочковидным формам. У кокков спорообразование происходит редко, а для вибрионов и спирилл оно несвойственно.
Процесс спорообразования заключается в том, что в определенном месте бактериальной клетки цитоплазма сгущается, затем этот участок покрывается плотной оболочкой. В течение нескольких часов бактериальная клетка превращается в спору.
Спора может располагаться в центре или на конце бактериальной клетки. Споры различных видов имеют неодинаковую форму. Они могут быть шаровидными, овальными. У некоторых видов их диаметр превышает толщину клетки, и это приводит к ее деформации – вздутию.
Спорообразование усиливается при наступлении неблагоприятных для развития условий, в частности, при обеднении питательной среды.
Споры более устойчивы, чем вегетативные формы этих же бактерий к действию проникающей радиации, ультразвука, высушивания, замораживания, разрежения, гидростатического давления, ядовитых веществ и др. Споры некоторых бактерий остаются жизнеспособными в течение 20 мин даже в кипящей концентрированной кислоте. Плотная, многослойная оболочка хорошо защищает споры от проникновения вредных веществ.
Благодаря способности к образованию спор, обладающих исключительно высокой устойчивостью к внешним воздействиям, спорообразующие бактерии остаются жизнеспособными при крайне неблагоприятных условиях.
Подавление жизнеспособности и уничтожение спорообразующих бактерий является одной из основных задач консервной промышленности, переработки и хранения сельскохозяйственных продуктов.
Спорообразование у бактерий не связано с размножением, так как бактериальная клетка способна образовывать лишь одну спору. Если споры попадают в благоприятные условия, то каждая из них в течение нескольких часов прорастает в вегетативную бактериальную клетку. Вначале лопается оболочка споры, а затем в этом месте появляется проросток клетки, постепенно превращающийся в нормальную клетку.
Нередко приходится наблюдать так называемые дремлющие споры. Споры, сохраняя жизнеспособность в течение долгого времени, могут прорастать постепенно через продолжительные сроки: от нескольких суток до многих лет.
1.2. Размножение бактерий
Существуют несколько способов размножения различных групп бактерий. Подавляющее число размножается путем деления клеток на две части (рис. 3).
Рисунок 3 – Деление бактериальной клетки
В средней части клетки образуется поперечная перегородка, которая разделяет клетку. Образовавшиеся новые клетки могут быть неодинаковыми по размеру. Скорость размножения зависит от условий питания, температуры, доступа воздуха и других факторов.
Размножению препятствуют истощение питательной среды, накопление продуктов обмена и другие физические, химические и биологические внешние факторы. Так, при снижении температуры на 10 °С скорость размножения снижается в 2–3 раза.
Попадая в новые условия, на свежий субстрат, микробы не сразу начинают размножаться. В течение некоторого времени они приспосабливаются к среде обитания, затем начинается бурное размножение, замедляющееся по мере исчерпания питательных ресурсов и накопления продуктов жизнедеятельности.
Быстрое развитие микробиологической порчи продуктов (окисления, гниения и др.) обусловлено исключительно высокой скоростью развития и размножения бактерий.
Кокки в процессе размножения делятся в одной, двух или трех взаимно перпендикулярных плоскостях. После деления они остаются слабо скрепленными друг с другом, в результате чего возникают сочетания кокков, отличающиеся по взаимному расположению (рис. 4): диплококки – парные кокки; стрептококки – цепочки кокков; тетракокки – по четыре кокка; capцины – в форме правильных пакетов по 8, 16 штук; стафиллококки – скопления, напоминающие гроздья винограда.
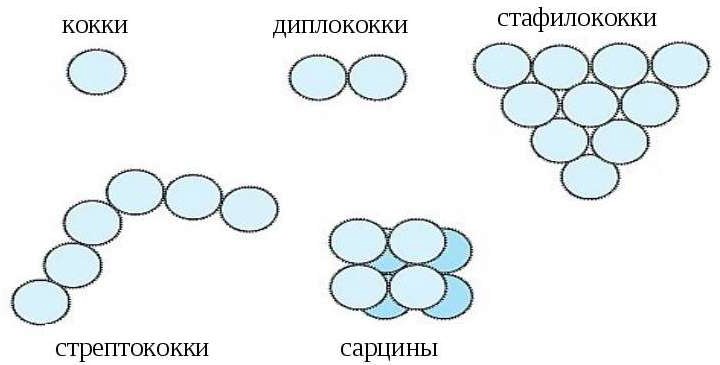
Рисунок 4 – Формы кокков
При очень слабой связи между возникающими при делении клетками образуются микрококки, во взаимном расположении которых нет никаких закономерностей. Они расположены поодиночке или в виде случайных скоплений по несколько экземпляров.
Палочки (бактерии, бациллы), подобно коккам, могут располагаться парами по длине – диплобактерии и цепочками – стрептобактерии. Большинство палочек располагается одиночно. Известны палочки строго цилиндрической формы, бочковидные, с резко обрубленными, вогнутыми или заостренными концами и др.
1.3. Систематика бактерий
Систематизация – упорядочение представлений о любых объектах и группах живых существ, в том числе микроорганизмов, необходима для облегчения распознавания этих объектов, установления степени родства или сходства между отдельными особями или группами особей. Систематизация микроорганизмов существенно облегчает практическую работу с ними.
В настоящее время в микробиологической практике все бактерии в зависимости от типичной формы клеток принято делить на семейства палочковидных, шаровидных и извитых.
Поскольку в каждое семейство объединяется множество разнообразных организмов, семейства подразделяют на роды. Так, семейство шаровидных бактерий в зависимости от характера объединения клеток в группы делят на роды микрококков, стрептококков, сарцин.
В семействе палочковидных бактерий различают два рода: род собственно бактерий, к которому относят все неспособные к образованию спор и род бацилл, объединяющий бактерии, которые способны образовывать споры.
Семейство извитых бактерий принято делить в зависимости от степени извитости на роды вибрионов (бактерии, изогнутые в виде запятой), спирилл и спирохет.
Деления на роды недостаточно для ориентировки в свойствах бактерий, поскольку оно основано только на внешних признаках. В каждый род объединяются бактерии, сходные по внешним признакам, но часто имеющие совершенно различные физиологические особенности и свойства.
В связи с этим роды делят на виды. Вид – систематическая категория, объединяющая организмы не только по внешним, но и по физиологическим признакам, а также по признакам родственного происхождения.
Для определения вида бактерий, кроме морфологических признаков (подвижность, отношение к диагностическим окраскам и т. д.), используются физиологические (потребность в кислороде, способность сбраживать различные сахара и т. д.), культуральные (характер образуемых колоний, особенности роста на некоторых питательных средах и т. д.) и др. Наименование вида бактерий состоит из двух слов, первое обозначает принадлежность к роду, а второе непосредственно указывает вид. Например, название «бактериум флуоресценс» означает, что микроорганизм относится к палочковидным бесспоровым бактериям (род бактерий), образующим пигмент флюоресцеин (вид – флуоресценс); «стрептококкус лактис» – относится к шаровидным бактериям, образующим цепочки из нескольких кокков (род стрептококков), способным вызывать скисание молока (вид – лактис).
1.4. Дыхание бактерий
Дыхание, или биологическое окисление, основано на окислительно-восстановительных реакциях, идущих с образованием АТФ-универсального аккумулятора химической энергии. Энергия необходима микробной клетке для ее жизнедеятельности. При дыхании происходят процессы окисления и восстановления: окисление – отдача донорами (молекулами или атомами) водорода или электронов; восстановление – присоединение водорода или электронов к акцептору. Акцептором водорода или электронов может быть молекулярный кислород (такое дыхание называется аэробным) или нитрат, сульфат, фумарат (такое дыхание называется анаэробным – нитратным, сульфатным, фумаратным).
Анаэробиоз (от греч. aer – воздух, bios – жизнь) – жизнедеятельность, протекающая при отсутствии свободного кислорода. Если донорами и акцепторами водорода являются органические соединения, то такой процесс называют брожением.
При брожении происходит ферментативное расщепление органических соединений, преимущественно углеводов, в анаэробных условиях. С учетом конечного продукта расщепления углеводов различают молочнокислое, спиртовое, уксусное и другие виды брожения.
1.5. Молочнокислое брожение
Гомоферментативное молочнокислое брожение. Начинается с гликолитического пути, 10 из 11 реакций у этих процессов идентичны. Основными субстратами служат моносахара (глюкоза) и дисахара (мальтоза, лактоза). Брожения более сложных субстратов проходят через путь предварительного расщепления их до глюкозы или продуктов ее превращения (глюкозо-6-фосфата). Конечный выход энергии на окислительном этапе распада глюкозы составляет 2 молекулы АТФ на 1 моль глюкозы. Помимо этого образуется 2 молекулы ПВК и 2 молекулы восстановленного переносчика (НАД-H2). Молекула ПВК – достаточно окисленное соединение и может служить акцептором электронов. В этом случае при участии фермента лактатдегидрогеназы 2 электрона переносятся с НАД-H2 на молекулу пировиноградной кислоты, что приводит к образованию молочной кислоты.
глюкоза + 2ФН + 2АДФ → 2 молочная кислота + 2АТФ + 2H2O.
1.6. Гетероферментативное молочнокислое брожение
Начинается с пентозофосфатного пути. Этот путь позволяет использовать в качестве энергетического материала не только гексозы, но и пентозы, а также синтезировать рибозы, необходимые для построения нуклеиновых кислот и других соединений. Конечные продукты брожения – C2- и C3-фрагменты: 3-ФГА и ацетилфосфат.
3-ФГА претерпевает ряд ферментативных превращений, идентичных таковым гликолиза, и через пируват превращается в молочную кислоту. Судьба двухуглеродного фрагмента различна: двухступенчатое восстановление ацетилфосфата приводит к накоплению в среде этанола; окислительный путь превращения ацетилфосфата завершается образованием уксусной кислоты.
В качестве конечных продуктов образуются молочная и уксусная кислоты, этиловый спирт, глицерин, CO2 и др. Образование уксусной кислоты из ацетилфосфата сопряжено с синтезом АТФ.
Если брожение идет с образованием этанола, то выход энергии равен 1 молекуле АТФ на молекулу сброженной глюкозы; если образуется уксусная кислота, то общий энергетический баланс процесса составляет 2 молекулы АТФ на молекулу глюкозы, т. е. такой же, как при гликолизе:
глюкоза + ФН + АДФ → лактат + АТФ + этанол + СО2;
глюкоза + 2ФН + 2АДФ + НАД+ → лактат + 2АТФ + ацетат + СО2+ НАДН2.
1.7. Молочнокислые бактерии
Гомоферментативное молочнокислое брожение осуществляют молочнокислые бактерии, относящиеся к родам Lactococcus и Pediococcus, а также некоторые представители рода Lactobacillus. Гетероферментативные молочнокислые бактерии Leuconostoc mesenteroides сбраживают глюкозу в молочную кислоту, этанол и CO2. У других гетероферментативных молочнокислых бактерий больший удельный вес занимает накопление уксусной кислоты: Lactobacillus fermentum, Lactobacillus brevis. Большинство молочнокислых бактерий имеют два пути сбраживания углеводов: гликолитический и пентозофосфатный. Сбраживание гексоз, как правило, протекает по гликолитическому пути, а пентоз – по окислительному пентозофосфатному. Это имеет место у представителей рода Lactobacillus: casei, plantarum, xylosis.
1.8. Спиртовое брожение
Процесс спиртового брожения идет по пути гликолиза до образования ПВК. Затем, в результате ее окислительного декарбоксилирования при участии ключевого фермента спиртового брожения пируватдекарбоксилазы образуется, уксусный альдегид. Особенность реакции заключается в ее полной необратимости. Образовавшийся ацетальдегид становится конечным акцептором водорода и восстанавливается до этанола с участием НАД+ – зависимой алкогольдегидрогеназы. В итоге из 1 молекулы гексозы образуются 2 молекулы этилового спирта и 2 молекулы углекислоты. Сбраживание 1 молекулы глюкозы приводит к образованию 2 молекул АТФ:
глюкоза + 2 ФН + 2АДФ → 2этанол + 2АТФ + 2СО2.
1.9. Формы брожения по Нейбергу
В присутствии бисульфита основным продуктом брожения будет глицерин. Оказалось, что бисульфит образует комплекс с ацетальдегидом, и последний не может больше функционировать как акцептор электронов. Следствием этого является передача электронов от НАД-H2 на фосфодиоксиацетон, восстановление его до 3-фосфоглицерина и дефосфорилирование, приводящее к образованию глицерина. Кроме глицерина в среде происходит накопление ацетальдегида (в комплексе с бисульфитом), этанола и CO2, но образование последних двух продуктов подавлено:
глюкоза + бисульфит → глицерол + ацетальдегидсульфит + СО2.
Спиртовое брожение протекает обычно при pH 3–6. Если его проводить в щелочной среде, например, в присутствии NaHCO3 или Na2HPO4, также происходит накопление глицерина. Оказалось, что в щелочных условиях ацетальдегид участвует в реакции дисмутации с образованием уксусной кислоты и этилового спирта. Акцептором электронов, как и в предыдущем случае, служит фосфодиоксиацетон, который преобразуется в глицерин:
2 глюкоза + Н2О → этанол + ацетат + 2 глицерол + 2СО2.
В условиях свободного доступа кислорода воздуха процесс спиртового брожения ингибируется и активируется дыхание – эффект Пастера. Микроорганизмы, осуществляющие спиртовое брожение: Saccharomyces, Schizosaccharomyces, Saccharomycodes, Mucor, Sarcina ventriculi, Erwinia amylovora, Zymomonas mobilis. В анаэробных условиях у высших растений также отмечено накопление этилового спирта.
1.10. Маслянокислое брожение
В маслянокислом брожении происходят реакции конденсации С2-соединений, в результате чего образуется С4 – акцепторная кислота. В качестве конечных продуктов в процессе брожения возникают соединения различной степени восстановленности. Характерным продуктом брожения является масляная кислота. Превращение глюкозы до пирувата осуществляется по гликолитическому пути. Ключевая реакция – разложение пирувата до ацетил-КоА и CO2. Путь, ведущий к синтезу масляной кислоты, начинается с реакции конденсации двух молекул ацетил-КоА. Он не связан с получением клеткой энергии, функция – акцептирование водорода, образовавшегося в процессе гликолиза.
Другое направление – превращение ацетил-КоА, ведущее к синтезу ацетата, именно с этим путем связано дополнительное получение энергии (при этом синтезируется молекула АТФ). Основным источником выделяемых при брожении газообразных продуктов (CO2 и H2) служит реакция окислительного декарбоксилирования пирувата. Выведение уравнения маслянокислого брожения и определение его энергетического выхода затруднительно из-за лабильности процесса, состоящего из двух основных ответвлений: одного – окислительного, ведущего к образованию ацетата и АТФ, другого – восстановительного. Количественное соотношение между обоими ответвлениями зависит от многих внешних факторов. В целом на 1 моль сбраживаемой глюкозы образуется 3,3 моля АТФ. Это наиболее высокий энергетический выход брожения из всех рассмотренных выше типов.
1.11. Маслянокислые бактерии
Осуществляют такой тип брожения бактерии, относящиеся к роду Clostridium. Некоторые клостридии наряду с кислотами накапливают в среде нейтральные продукты (бутиловый, изопропиловый, этиловый спирты, ацетон). Это дало основание выделить как вариант маслянокислого брожения ацетонобутиловое брожение. У клостридиев, осуществляющих ацетонобутиловое брожение, образование масляной кислоты происходит на первом этапе брожения. По мере подкисления среды (до pH ниже 5) и повышения в ней концентрации жирных кислот индуцируется синтез ферментов, приводящих к накоплению нейтральных продуктов, в первую очередь н-бутанола и ацетона. Изучение физиологии группы клостридиев, осуществляющих ацетонобутиловое брожение, привело к открытию В.Н. Шапошниковым явления двухфазности этого процесса, которое позднее было обнаружено в большинстве типов брожений, характеризующихся сложным набором конечных продуктов.
По отношению к молекулярному кислороду бактерии можно разделить на три основные группы: облигатные, т. е. обязательные аэробы, облигатные анаэробы и факультативные анаэробы.
Облигатные аэробы – это прокариоты, для роста которых необходим кислород. К ним относят большинство прокариотных организмов.
Облигатные анаэробы (клостридии ботулизма, газовой гангрены, столбняка, бактероиды и др.) растут только на среде без кислорода, который для них токсичен. При наличии кислорода бактерии образуют перекисные радикалы кислорода, в том числе перекись водорода и супероксид-анион кислорода, токсичные для облигатных анаэробных бактерий, поскольку они не образуют соответствующие инактивирующие ферменты. Аэробные бактерии инактивируют перекись водорода и супероксид-анион кислорода соответствующими ферментами (каталазой, пероксидазой и супероксиддисмутазой).
Факультативные анаэробы могут расти как при наличии, так и при отсутствии кислорода, поскольку они способны переключаться с дыхания в присутствии молекулярного кислорода на брожение в его отсутствие. Они способны осуществлять анаэробное дыхание, называемое нитратным: нитрат, являющийся акцептором водорода, восстанавливается до молекулярного азота и аммиака.
Среди облигатных анаэробов различают аэротолерантные бактерии, которые сохраняются при наличии молекулярного кислорода, но не используют его. Для выращивания анаэробов в бактериологических лабораториях применяют анаэростаты – специальные емкости, в которых воздух заменяется смесью газов, не содержащих кислорода. Воздух можно удалять из питательных сред путем кипячения, с помощью химических адсорбентов кислорода, помещаемых в анаэростаты или другие емкости с посевами.
2. Грибы
Актиномицеты, или лучистые грибы – микроорганизмы близкие к бактериям. В значительных количествах они встречаются в почве в природных водных источниках. Это одноклеточные организмы с ветвистым строением тела. Нити ветвления, из которых состоит тело, называют мицелием. Часть мицелия развивается над поверхностью питательного субстрата.
Актиномицеты малотребовательны к условиям обитания по сравнению с другими микроорганизмами. Среди актиномицетов многие являются антагонистами болезнетворных бактерий. Они являются продуцентами некоторых антибиотиков. Размножаются актиномицеты с помощью спор, образующихся в большом количестве на воздушной части мицелия. Некоторые виды размножаются делением или перешнуровыванием клетки во многих местах.
При развитии на пищевых продуктах актиномицеты придают им землистый запах. Актиномицеты можно рассматривать как переходную форму между бактериями и более сложно организованными плесневыми грибами.
2.1. Плесневые грибы
Плесневые грибы, как и бактерии, относят к низшим споровым растениям, лишенным хлорофилла. Для своей жизнедеятельности грибы нуждаются в готовых органических веществах в связи с неспособностью самостоятельно образовывать органические вещества из углекислого газа. Нуждаются они и в доступе воздуха, так как без него развиваться не могут.
По строению клетки плесневые грибы принципиально не отличаются от клеток бактерий и дрожжей, но имеют одно, а иногда и несколько дифференцированных ядер. В цитоплазме их клеток часто образуются одна или несколько полостей – вакуолей, заполненных клеточной жидкостью. Образование вакуолей обусловлено старением белковых коллоидов цитоплазмы и снижением в связи с этим способности удерживать воду в связанном состоянии. Вакуоли являются резервуаром, собирающим вредные для клеток продукты жизнедеятельности.
Клетки имеют сильно вытянутую форму и поэтому напоминают нити, называемые гифами. Толщина их – 1–15 мкм. Они сильно ветвятся, образуя переплетающуюся массу – мицелий, или грибницу. Мицелий является телом плесневых грибов. Среди плесневых грибов встречаются одноклеточные и многоклеточные. Плесневые грибы широко распространены в природе.
Развиваясь на питательных субстратах, они образуют пушистые налеты различного цвета. Потребляя вещества субстрата, грибы производят глубокие изменения всех его составных частей, выделяют в него продукты своей жизнедеятельности. От этого пищевые продукты приобретают специфичные плесневые запах и вкус становятся ядовитыми. Использование пищи, пораженной плесневыми грибами, может вызвать тяжелые заболевания.
Характерной особенностью является способность плесневых грибов развиваться при низкой влажности субстрата (около 15 %), в связи с чем они могут поражать сухофрукты, сухари, а из непищевых товаров – бумагу, кожу, пряжу и ткани, прочность которых от этого значительно снижается.
Размножение плесневых грибов. Грибы способны размножаться многими способами.
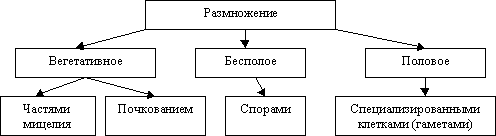
Рисунок 5 – Размножение грибов
Наиболее простым является размножение частями мицелия. Каждая часть мицелия, попав на новый участок субстрата, при благоприятных условиях становится самостоятельной и развивается как целый организм.
Часть мицелия, находящаяся в питательном субстрате, играет основную роль в обеспечении организма питательными веществами, влагой и др. Часть мицелия, поднимающаяся над поверхностью субстрата служит для образования конидий, спор, оидий, с помощью которых плесневые грибы размножаются.
Споры – тельца различной формы размерами до нескольких микрон. Они образуются из концах гиф воздушной части мицелия внутри особых образований овальной и полукруглой формы – спорангий. Спорангиоспоры образуются путем распадения многоядерной протоплазмы молодого спорангия на множество отдельных участков, которые постепенно покрываются собственной оболочкой и превращаются в споры.
Нити воздушного мицелия, несущие спорангии, называются спорангиеносцами. Такое образование спор характерно для одноклеточных грибов. У многоклеточных формируются так называемые экзоспоры, т. е. внешние или наружные, которые чаще называются конидиями, а воздушные гифы, несущие их, конидиеносцами.
Конидии образуются путем отделения непосредственно от конидиеносца или от особых клеток, расположенных на его вершине. Эти клетки обычно имеют продолговатую форму и называются стеригмами. Конидии располагаются на конидиеносцах поодиночке, цепочками.
Спорангиеносцы и конидеоносцы образуют видимый пушистый налет на поверхностях материалов, пораженных грибами. Различная окраска налета (зеленая, оливковая, розовая, белая, серая и др.) зависит от окраски конидий, спор, оидий. Мицелий грибов бесцветен.
Грибы, способные размножаться половым путем, называют совершенными.
Грибы, которые никогда не размножаются половым путем, относят к несовершенным.
Многие грибы способны при наступлении неблагоприятных условий образовывать покоящиеся стадии в виде так называемых склероций. Это крепкие, твердые, с поверхности темные, а внутри белые желвачки различных размеров и форм, образованные из плотно переплетенных гиф. Склероции, попадая в благоприятные условия, прорастают и образуют обычно те или иные (в зависимости от вида гриба) органы размножения. Они часто образуются в колосьях злаков. Другой покоящейся стадией являются хламидоспоры. При их образовании цитоплазма внутри гиф собирается в виде комочков, образует новую оболочку, обычно толстую и окрашенную, и гифы становятся похожими на цепочки или четки, состоящие из хламидоспор. Иногда хламидоспоры образуются только на концах гиф (рис. 6).
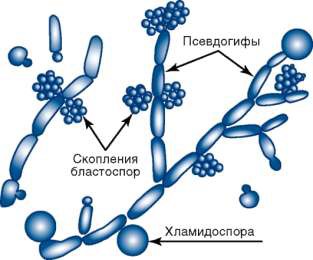
Рисунок 6 – Хламидоспоры
Систематика грибов. В зависимости от особенностей строения мицелия, строения органов бесполого размножения, способности к половому размножению и характера плодовых тел все грибы делят на пять классов.
Архимицеты – грибы, развивающиеся без образования мицелия или образующие слабо развитый мицелий. Размножаются преимущественно бесполым путем, образуя жгутиковые подвижные споры. Это наиболее примитивные грибы. Большинство представителей этого класса являются паразитами высших форм живых организмов. Гриб Olpidium brassicae (ольпидий капустный) вызывает заболевание капустной рассады, поражающее шейку стебля («черная ножка»), Synchуtrium endobiоticum вызывает бугристость клубней картофеля (рак картофеля), поражая также молодые побеги и столоны клубней.
Фикомицеты – имеют хорошо развитый одноклеточный мицелий. Размножение половое и бесполое. Споры находятся в спорангиях. Многие грибы этого класса вызывают заболевания растений. Фитофтора поражает клубни и ботву картофеля, плоды томатов, баклажаны. На клубнях, зараженных фитофторой, образуются вдавленные пятна, захватывающие постепенно весь клубень, который при этом отмирает. Поверхность клубня покрывается белым пушистым налетом, состоящим из спорангиеносцев. Мицелий гриба и споры сохраняются на клубнях, на остатках ботвы, в почве.
Другой представитель этого класса – Plasmopara – вызывает заболевание винограда, поражающее листья и ягоды. Побуревшие листья опадают, ягоды сморщиваются, часто теряют способность к созреванию и осыпаются. Споры гриба сохраняются на растительных остатках в почве в течение нескольких лет. Mucor, часто именуемый «серой плесенью», является исключительно неприхотливым, быстро развивающимся возбудителем порчи многих продуктов – плодов, овощей, кулинарных изделий и других товаров.
Аскомицеты – сумчатые грибы с ветвистым многоклеточным мицелием.
Класс аскомицетов насчитывает большое количество видов, различных по строению и свойствам. Среди них много паразитов культурных растений, возбудителей порчи пищевых продуктов.
Размножаются аскомицеты бесполым путем с помощью конидий. При половом размножении этих грибов образуются споры в особых сумках (асках). Некоторые сумчатые грибы не имеют плодовых тел, и сумки у них развиваются непосредственно из мицелия. Грибы, образующие плодовые тела, называются плодосумчатыми, а не образующие плодовые тела – голосумчатыми.
Примером голосумчатых грибов могут служить дрожжи и дрожжеподобные грибы, например Endomuces vernalis, способный, развиваясь на самых дешевых субстратах, накапливать до 30 % жира. В связи с этим он применяется в промышленности.
Широко распространенными плодосумчатыми грибами являются грибы родов Aspergillus и Penicillium. Плодовые тела у них в виде мелких шариков, внутри которых находятся сумки со спорами. Конидии отдельных видов различаются по форме и окраске, строение конидиеносцев также разнообразно.
Грибы рода Aspergillus имеют одноклеточные и неразветвленные конидиеносцы. Верхушки конидиеносцев образуют утолщение, несущее на поверхности бутылковидные стеригмы с цепочками конидий.
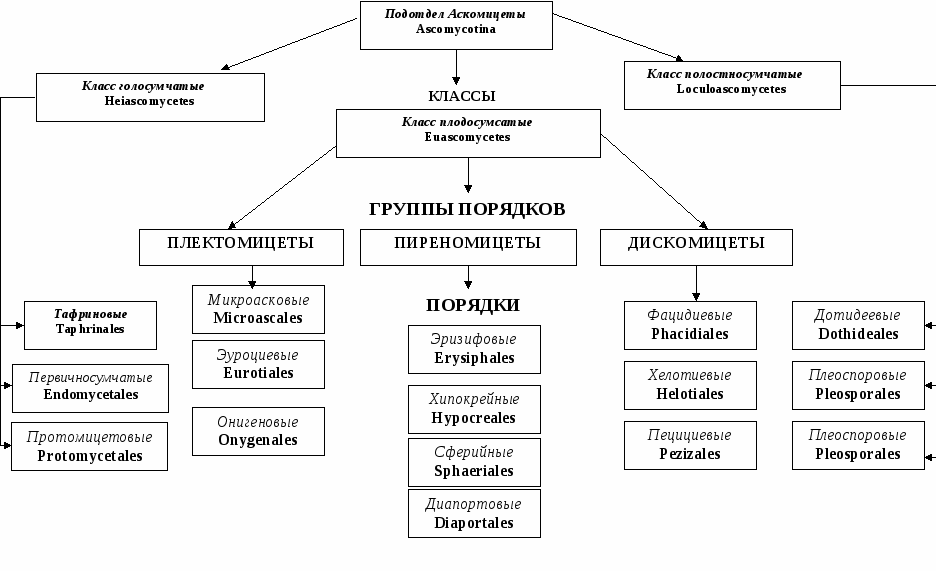
Рисунок 7 – Систематика подотдела сумчатые грибы
У грибов рода Penicillium конидиеносцы многоклеточные, ветвящиеся. На концах разветвлений находятся стеригмы с цепочками конидий. Эта группа совершенных многоклеточных грибов наиболее широко распространена в природе в силу большой устойчивости к неблагоприятным условиям окружающей среды. Прорастают грибы в виде сначала белого, а потом матово-зеленого налета почти на всех товарах и материалах, имеющих достаточную влажность, являются возбудителями плесневения пищевых продуктов и различных органических материалов, например, обоев, кожи, хлопкового волокна, клея, тканей и др. Отдельные виды родов Aspergillus и Penicillium вызывают заболевания (микозы) у человека и животных.
Некоторые виды грибов этого класса широко используют в технологиях. Aspergillus применяют в производстве лимонной кислоты из сахара, для осоложения зерна, в промышленном производстве спирта. Некоторые виды Penicillium выращивают для получения лечебного препарата пенициллина, другие играют важную роль в созревании сыра рокфор.
Гриб этого же класса Sclerotinia является активным возбудителем порчи плодов и овощей – огурцов, моркови (белая гниль моркови).
Спорынья – паразит злаковых растений (ржи, пшеницы, ячменя), вызывающий их заболевание под таким же названием. В колосьях растений появляются темно-фиолетовые рожки, представляющие склероции гриба.
Basidiomycota имеют ветвистый многоклеточный мицелий. Размножение половое и бесполое. Представлены в природе также большим числом разновидностей. К этому классу относят все известные шляпочные грибы, трутовики, домовые грибы. Многие шляпочные грибы имеют существенное хозяйственное значение в связи с использованием в пищевых целях. Трутовики являются опасными разрушителями живой древесины, деревянных строительных материалов, имеющих повышенную влажность. Домовые грибы – специфичные возбудители порчи мертвой древесины. Они способны разрушать деревянные конструкции, имеющие даже невысокую влажность.
К этому же классу относят большое число паразитических грибов, поражающих важные в хозяйственном отношении растения: плодовые деревья, ягодники, огородные и полевые культуры. Наиболее известны головневые и ржавчинные грибы. Головневые грибы паразитируют на злаковых растениях, вызывают различные их заболевания – мокрую головню, пыльную головню, пузырчатую головню (на кукурузе). Растения, пораженные этими грибами, становятся черными, словно обожженными. Попадание спор головни в муку придает ей неприятный селедочный запах, который сохраняется в хлебе. Споры головни вызывают расстройства кишечника, раздражение слизистых желез и другие болезненные явления. Пораженность головней продовольственного зерна нормируется и не должна быть более 5 %.
Несовершенные грибы имеют многоклеточный мицелий. Размножаются конидиями и оидиями. Половым путем не размножаются. Несовершенные грибы широко распространены в природе. Многие из них вызывают плесновение пищевых продуктов и различных материалов (тканей, хлопка, шерсти, бумаги и др.). Другие грибы паразитируют на культурных растениях.
Наиболее важным с хозяйственной точки зрения являются следующие роды этих грибов.
Fusarium вызывает заболевание картофеля, называемое сухой гнилью. Пораженные участки клубня сморщиваются, превращаются в сухую крахмалистую массу. Этот гриб часто поражает корни бобовых, тыквенных и других растений, вследствие чего последние быстро засыхают. Некоторые виды относятся к активным возбудителям болезней луковичных растений, особенно цветочных. Другие, развиваясь на злаках, делают зерна ядовитыми. Все эти заболевания известны под названием «фузариозы».
Botrytis вызывает шейковую гниль лука, серую гниль различных овощей (капусты, моркови, помидоров и др.) и ягод (малины, земляники, крыжовника и др.). Пораженные овощи и ягоды покрываются пушистым серым налетом, ткани их становятся водянистыми, буреют, размягчаются.
Alternaria поражает корнеплоды в период хранения, вызывая черную гниль. На корнеплодах появляются сухие вдавленные темносерые пятна.
Oidium образует сильно разветвленный мицелий, который у взрослых грибов распадается на оидии. Грибы этого рода часто развиваются в виде белой или кремово-белой бархатистой пленки на поверхности сметаны, кисломолочных и других продуктов, могут поражать также сливочное масло. Развиваясь в поверхностном слое квашеных овощей, опресняют его, используя молочную кислоту и превращая ее в воду и углекислый газ. Опресненный слой становится доступным для гнилостых микроорганизмов, которые и вызывают порчу этих продуктов.
Monilia портит семечковые и косточковые плоды, вызывая образование бурых пятен и размягчение тканей. Разрушает плоды во время роста и созревания и продолжает развиваться на хранящихся плодах. Конидии гриба сохраняются в почве и мумифицированных грибом плодах. Поверхность таких плодов черного цвета, блестящая; они прочно удерживаются на ветвях дерева.
3. Дрожжи
Дрожжи относят к классу сумчатых грибов. Выделение их в отдельную группу объясняется исключительно широким применением дрожжей в ряде производств – хлебопечении, виноделии, производстве спирта, пивоварении и др. Большинство дрожжей способно превращать различные углеводы в этиловый спирт, углекислый газ, на чем и основывается их использование.
Дрожжи применяются как пищевой и кормовой продукт в связи со способностью накапливать в клетках большие количества хорошо усвояемых организмом человека белков, жиров, разнообразных витаминов, минеральных веществ.
Клетки дрожжей имеют эллипсовидную или яйцевидную форму.
Встречаются круглые, цилиндрические.
Размеры дрожжевых клеток довольно велики по сравнению с бактериями. В зависимости от вида дрожжей, их возраста и условий питания диаметр дрожжевых клеток 1–5 мкм, длина до 15 мкм.
Строение дрожжей. В каждой дрожжевой клетке имеется четко отграниченное от цитоплазмы ядро. Оно хорошо различимо при обычной микроскопии в препаратах, окрашенных специальными (ядерными) красителями, связывающимися с веществами ядра и не окрашивающими цитоплазму (рис. 8).
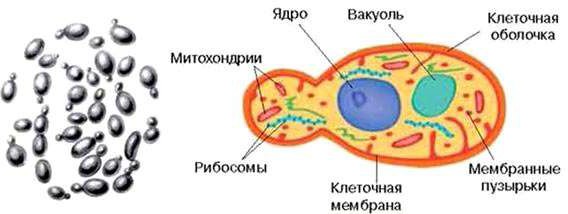
Рисунок 8 – Строение дрожжевой клетки
У всех дрожжей отсутствуют жгутики, поэтому они неподвижны. В цитоплазме дрожжей имеются крупные вакуоли, а также тельца, аналогичные по функциям мезосомам, – митохондрии.
3.1. Размножение дрожжей
Различные представители этой группы микроорганизмов размножаются по-разному. Большинство дрожжей размножается почкованием. При этом на теле клетки образуется одно, а иногда несколько вздутий, в которые из материнской клетки поступает часть ядра, цитоплазмы и прочие структурные элементы. Затем почка отделяется от клетки. Иногда почки, еще не отделившись, начинают в свою очередь почковаться. В результате образуется целое скопление дрожжевых клеток (сросток почкования).
У некоторых видов дрожжей размножение осуществляется делением. Образуется перегородка, в результате чего клетка делится на две, не всегда равные части. При последовательном делении может образоваться целая цепочка, которая в конечном итоге распадается на отдельные дрожжевые клетки.
Размножение дрожжей – процесс более длительный, чем у бактерий (рис. 9). В благоприятных условиях он происходит в течение нескольких часов.
Некоторые дрожжи, развиваясь на поверхности жидких субстратов, образуют плотные морщинистые пленки, под которыми субстрат подвергается порче или его вещества превращаются в углекислый газ и воду, в результате чего снижается выход полезных продуктов. Особенно большой ущерб от таких дрожжей в виноделии, пивоварении, при хранении квашеных овощей в дошниках.
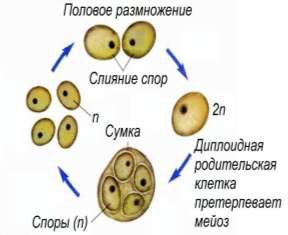
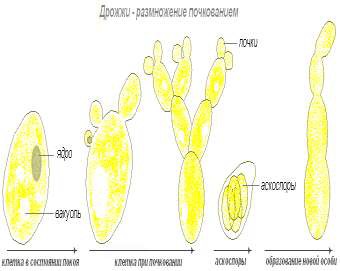
Рисунок 9 – Размножение дрожжей
Многие дрожжи размножаются спорообразованием. Количество спор в дрожжевой клетке от 1 до 12, но чаще четыре. Споры имеют круглую, овальную и другие формы. Образование спор может происходить бесполым и половым путями.
Споры дрожжей более устойчивы к вредным воздействиям, чем вегетативные клетки. Спорообразование наступает после периода вегетативного размножения, т. е, после некоторых изменений в составе среды, побуждающих к спорообразованию. Неблагоприятные условия, в частности голодание, также способствуют образованию спор.
3.2. Классификация дрожжей
Дрожжи классифицируются в зависимости от особенностей размножения, способности или неспособности к использованию некоторых сахаров и других признаков.
По способности к образованию спор дрожжи принято делить на два семейства: настоящие (сахаромицеты) и ложные (несахаромицеты).
К настоящим относятся дрожжи, которые могут размножаться как вегетативным способом, так и спорами. При этом материнскую клетку, в которой образуются споры, можно рассматривать как аскус (сумку), а споры можно называть аскоспорами.
К ложным дрожжам относятся дрожжи, которые не способны размножаться спорами. Семейства делятся на роды.
Большинство используемых человеком настоящих дрожжей (культурных) относится к роду Saccharomyces. Дрожжи этого рода размножаются почкованием, способны к бесполому образованию спор. Виды этого рода различаются особенностями вызываемого ими брожения, соотношением различных конечных продуктов при сбраживании, отношением к температуре, способностью сбраживать различные сахара.
Разновидности дрожжей, которые различают по признакам и называются расами, возникшим в результате многовекового культивирования в искусственных условиях.
В спиртовой промышленности часто применяются расы, способные быстро и полно сбраживать сахар при температуре 28–30 °С и устойчивые к действию спирта. В пивоварении используют расы, медленно развивающиеся и способные к брожению при низких температурах (4–10 ºС) и позволяющие получать хорошо осветляющееся ароматное пиво с низким содержанием спирта. Особые расы дрожжей с высокой подъемной силой применяют в хлебопекарном производстве.
Из семейства ложных дрожжей широко распространены в природе представители рода Torulopsis. Виды этого рода способны вызывать слабое спиртовое брожение, в связи с чем их используют в заквасках для кефира, кумыса. Их выращивают в больших количествах на дешевых материалах, даже на некоторых фракциях нефти как кормовые. Добавка дрожжей в корм животных увеличивает привесы скота, позволяет удешевлять откорм.
Род Mycoderma образует морщинистые прочные пленки на поверхности квашеных овощей, в чанах, где ведется сбраживание пивного сусла или сахарных растворов при производстве уксуса. Вред, приносимый этими дрожжами, заключается в быстром использовании субстратов за счет глубокого окисления сахаров (до углекислого газа и воды) и в придании готовому продукту несвойственных вкуса и запаха.
Многие дрожжи, развиваясь на пищевых продуктах, образуют на них налеты или колонии различной окраски. Некоторые способны вызывать тяжелые заболевания человека, поражая слизистые покровы, центральную нервную систему.
4. Вирусы
Существует множество инфекционных заболеваний человека, животных и растений, возбудители которых невидимы в оптическом микроскопе.
Л. Пастер предположил, что причиной таких заболеваний являются микроорганизмы, размеры которых много меньше размеров бактерий. На существование таких мельчайших микроорганизмов указывал и Н.Ф. Гамалея. Существование невидимых в микроскоп возбудителей болезней было окончательно установлено в 1892 г. русским ученым Д.И. Ивановским. Эти микроорганизмы являются вирусами.
Размер вирусов исчисляется в нанометрах. Вирусы свободно проходят через бактериальные фильтры, поры которых чрезвычайно мелки и не пропускают обычные бактерии. Поэтому они и названы фильтрующимися вирусами. Величина некоторых вирусов всего в несколько раз превышает величину крупных белковых молекул. Например, частицы вируса ящура имеют размер 8–12 нм. Вирусами средних размеров являются возбудитель гриппа (80–120 нм), бешенства (100–150 нм). Один из наиболее крупных вирусов – вирус натуральной оспы – имеет размер 120–200 нм.
Частицы различных вирусов имеют определенную форму: шарообразную, прямоугольную, нитевидную. Вирусы (рис. 10) можно видеть лишь с помощью микроскопов, дающих увеличение в десятки тысяч раз (электронных и др.).
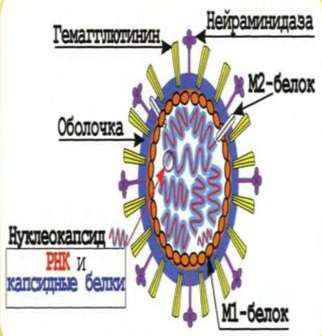
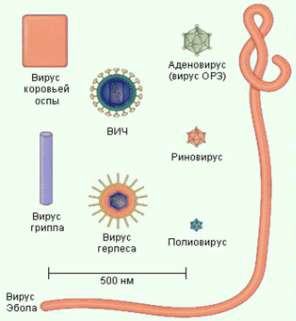
Рисунок 10 – Морфология вирусов
Важнейшим отличием вирусов от бактерий является их неспособность размножаться вне организма, поэтому вирусы не растут на обычных питательных средах, на которых хорошо развиваются бактерии и грибы. Вирусы являются внутриклеточными паразитами и размножаются только в живых клетках. Вызывая заболевания растений (табачная мозаика, карликовость томатов, парша картофеля и др.) и животных, вирусы наносят громадный ущерб народному хозяйству. Устойчивость вирусов к некоторым внешним воздействиям, например, к высокой температуре, не очень высока. Практически все они погибают при непродолжительном кипячении. Действие высушивания и низких температур переносят сравнительно легко.
Другой разновидностью ультрамикробов являются бактериофаги (пожиратели бактерий). Бактериофаги – это специфичные паразиты обычных бактерий. Проникая в клетки бактерий, бактериофаги вызывают их лизис, т. е. растворение. Явление бактериофагии впервые наблюдал Н.Ф. Гамалея в опытах с культурой сибиреязвенных палочек. В настоящее время известно множество бактериофагов, каждый из которых способен ликвидировать лишь определенные виды бактерий. Это свойство позволяет использовать фаги при распознавании выделенных при анализах культур.
Бактериофаг действует на живые, активно размножающиеся бактерии. Мертвые бактериальные клетки бактериофаг не растворяет. В некоторых случаях бактерии, оставшиеся живыми в присутствии бактериофага, приобретают к нему устойчивость.
Явление бактериофагии возникает на пищевых производствах, связанных с использованием микроорганизмов. Фаги растворяют микробы, участвующие в технологическом процессе (брожении, сквашивании), нанося этим ущерб производству.
Бактериофаги применяют в качестве профилактических и лечебных средств для борьбы с некоторыми инфекционными заболеваниями: дизентерией, брюшным тифом, холерой, дифтерией и др. В настоящее время известны фаги, паразитирующие не только на бактериях, но и на актиномицетах (актинофаги), плесневых грибах (микофаги).
Ряд признаков – прохождение через бактериальные фильтры, неспособность развиваться на мертвых органических субстратах – позволяют считать бактериофагов разновидностью вирусов.

